
Title of paper under discussion
Cortical Regions Involved in the Generation of Musical Structures during Improvisation in Pianists
Authors
Sara L. Bengtsson, Mihály CsÍkszentmihályi and Fredrik Ullén
Journal
Journal of Cognitive Neuroscience, vol 19, no 5, pp. 830–842
Link to paper (free access)

Overview
Scan the brain of someone in the act of selecting freely, for example moving a hand hand at will or choosing any number from 1 to 10, and a couple of regions will typically “light up”: the ‘dorsolateral prefrontal cortex’ and ‘rostral premotor cortex’. Would this be the same for an improvising musician?
Using functional magnetic resonance imaging (fMRI), Bengtsson and her colleagues scanned the brains of 11 professional pianists as they improvised on a simple prescribed melody, a task called ‘Improvise’. The pianists were then asked to repeat that same improvisation as accurately as possible from memory, a task called ‘Reproduce’. Finally the pianist was asked to do a free improvisation, with no need to memorise it, a task called ‘FreeImp’.
The scientists reasoned that by contrasting the ‘Improvise’ scan with the ‘Reproduce’ one they might reveal those regions of brain that were active due to improvisation, as the contrast would delete brain activity present in both tasks (eg pianistic movements and sensations). In case the ‘Improvise’ scans were also picking up ‘memorising’ brain activity needed for that task, the researchers compared their ‘Improvise vs Reproduce’ contrasts with ‘FreeImp vs Resting state’ contrasts. Brain areas active in both contrasts, they argued, must be underpinning the act of musical improvisation.
As predicted, such activated brain regions did include the right ‘dorsolateral prefrontal cortex’ and ‘rostral premotor areas’, plus the posterior part of the left ‘superior temporal gyrus’. The authors suggest these regions “are part of a network involved in musical creation”.
Method
Eleven Swedish male right-handed concert pianists volunteered for the study. Though lying in an fMRI scanner throughout the experiment, each participant was able to ‘play the piano’ (a one-octave piano midi keyboard) and read music (from a screen via a binocular/mirror system).
The experiment was composed of four parts, or ‘conditions’: Improvise, Reproduce, Rest and FreeImp. In all conditions a musical score consisting of a simple eight-bar musical melody was displayed, for example:

After 8 seconds of silently reading the score the participant was given instructions according to the experimental condition.
For Improvise, they were asked to use the displayed melody as a basis for an improvisation. The instructions to the participants were to “employ any kind of modifications of the presented melodic template they wished […]They were also instructed to memorize the performance for subsequent reproduction”
For Reproduce, again using the keyboard, “the task was to reproduce, as faithfully as possible, the improvisation previously made upon the same melody.”
For Rest, the instructions were to remain “relaxed, viewing the screen without any movements.”
For FreeImp, “the instruction was to improvise on the melody, as in Improvise, but without trying to memorize the performance.”
The experiment was repeated 9 times, with a different melody used each time.
As well as constantly monitoring each participant’s brain activity, the experimenters also made a careful midi-recording of each of their piano performances; this enabled them, for example, to record how accurately the pianist had managed to reproduce a given ‘Improvise’ improvisation in the subsequent Reproduce condition, and to compare FreeImp complexity with Improvise complexity.
Results
Styles of improvisation
Looking across all the improvisation recordings, Bengtsson and her colleagues counted up 11 different types of musical device used by the pianists to elaborate or modify their given melodies (or ‘templates’):
“i) Insertion of a fast group of one or more grace notes before a template note; (ii) substitution of a template note for another note; (iii) figuration, that is, expansion of the original template into melodic figures; (iv) insertion of a trill on a template note; (v) filling in, that is, insertion of chromatic or diatonic scales between template notes; (vi) repetitions of template notes; (vii) elimination of template notes; (viii) insertion of figures giving a broken two-part polyphony; (ix) rhythmization of the template; (x) insertion of a tremolo, that is, a trill-like figure between two notes with a larger interval than a second; and finally, (xi) switching of tonality from major to minor.” Examples of (i) to (v) are shown here:

Accuracy of the Reproduce performances
To give their fMRI analysis credence, the scientists needed to establish whether the Reproduce performances were similar enough in execution to the Improvise performances they were supposed to mimic. If so, then their proposal – that any contrast between an Improvise scan and its respective Reproduce scan would reveal the brain activity involved in the act of improvisation – should be sound. If not, then such a contrast may also be showing up brain activity due to the differences in physical execution between the two types of performance.
Comparing Improvise performances with Reproduce performances, the scientists found there was no significant difference in the number of notes played, nor in performance duration. To further investigate how well each improvisation was reproduced the researchers carried out a note-by-note comparison between each Improvise performance and its subsequent Reproduce performance, assigning each comparison an accuracy score called ‘LED’ (‘Levenshtein edit distance’). They found that “the participants were able to reproduce their performances in Improvise with remarkable accuracy in the Reproduce condition.”
FMRI scan contrasts
Having established that the pianists were remarkably accurate in their Reproduce performances, the scientists could move on to the analysis of brain imagery confident that the Improvise-Reproduce contrasts would indeed reveal brain activity solely involved in the act of improvisation (though perhaps also including brain activity associated with the act of memorising).
Four brain regions were identified as being more active during Improvise than Reproduce: the right Dorsolateral Prefrontal Cortex (DLPFC), Rostral Premotor Cortex (RPC), left Superior Temporal Gyrus (STG) and the middle occipital gyri (MOG):
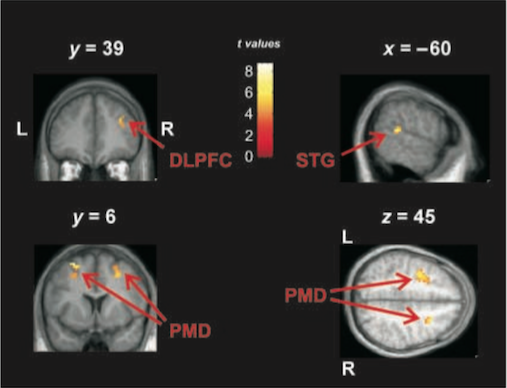
in order to establish that these brain regions were ‘lighting up’ due to the act of improvisation and not memorising, the researchers also contrasted the scans taken during the FreeImp condition with those of the Rest condition. They reasoned that any areas active in the FreeImp-Rest contrast that were also active in the Improvise-Reproduce contrast must be solely involved in improvisation, and not also memorisation, because during FreeImp the instruction was not to memorise the performance.
It turned out that the all these four brain regions, or areas very close to them, were also active in the FreeImp-Rest contrast, leading the researchers to assert with more confidence that the right DLPFC, the right and left RPC, the left STG and the right and left MOG are all critical for the act of musical improvisation. In the words of the authors “we therefore argue that the activity in these brain regions reflects neural processes involved in the generation of new musical material during improvisation.”
Another piece of analysis further implicated a specific sub-region of RPC, the right pre-Supplementary Motor Area (right pre-SMA), as being especially involved in the act of improvisation. By comparing the complexity of each Improvisation with that performer’s right pre-SMA brain activity during its execution, the scientists discovered that the more complex the improvisation, the more active was the right pre-SMA. In other words, the harder the pianist was improvising, the harder the right pre-SMA was working.
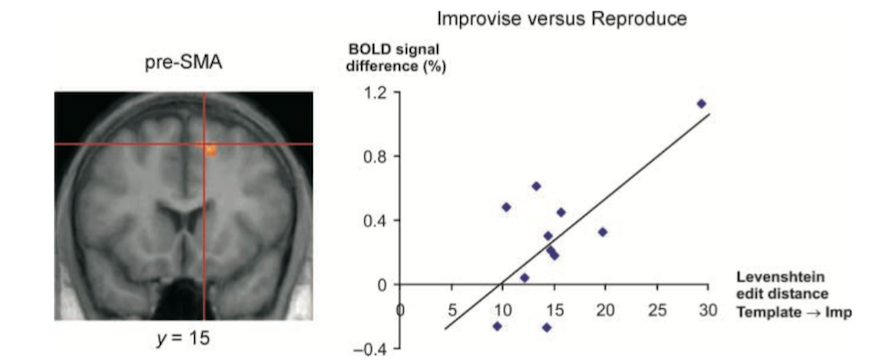
In a similar vein, the DLPFC was generally more active during FreeImp than Improvise, which could be related to the slightly higher level of improvisational complexity seen in Freeimp compared with Improvise.
Discussion
What is it about these particular brains regions that makes them ‘light up’ during improvisations?
The authors looked at the findings of other brain researchers in an effort to ascribe specific improvisational roles to each of these regions.
Dorsolateral Prefrontal Cortex (DLPFC)
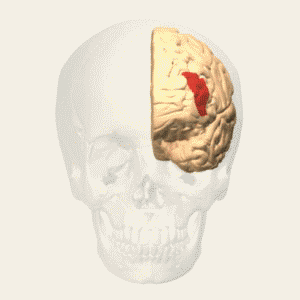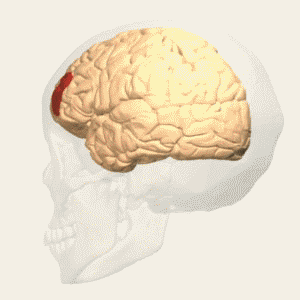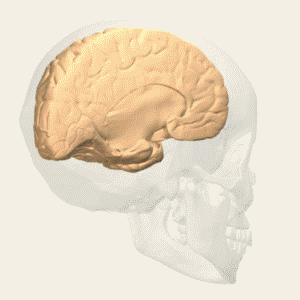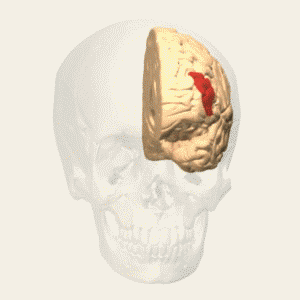
Previous research papers on many different types of motor task have consistently implicated the DLPFC when movement is freely chosen compared with movement in ‘repetitive or externally determined” tasks. Similarly, the authors explained, DLPFC acitivity is related to free selection in cognitive tasks, such as word generation or number generation.
Some research suggests that the DLPFC is especially involved in paying attention to free choice selection, while other research implicates it in the actual selection process itself. Memory researchers have suggested that the DLPFC is implicated in inhibiting stereotyped, habitual responses in favour of random (or at least pseudo-random) ones, achieving this by constantly monitoring such responses in ‘working memory’.
In addition to these specific roles – attention to action, monitoring in working memory, response selection and suppression of stereotype responses – the authors claim that their results are consistent with “a vast body of neurological and experimental data” claiming a “central role for the DLPFC in planning and performance of novel or complex behavioural sequences, including language and thought”. Thus, claims Bengtsson, the DLPFC has “one central function” during improvisation, which is to be “supervisory: to maintain and execute an overall plan for the improvisation through top-down influences on the activity”, with the RPC areas – with which it has rich connections – subordinate to its control. In this way “a whole set of freely selected modifications of the original melody” can be organised in time “according to a musically meaningful overall plan”.
Rostral premotor cortex (RPC)
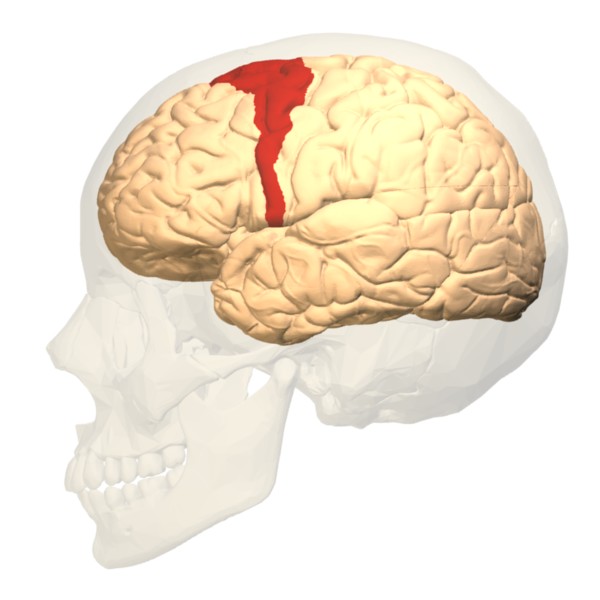
Having suggested that the RPC is subordinate to the DLPFC in the act of improvisation, our authors identify two sub-areas of RPC – the pre-Supplementary Motor Cortex (pre-SMA) and the Dorsal Premotor Cortex (PMD) – that may each support different improvisatory roles.
The pre-SMA, according to previous research, is especially involved in timing and rhythmic sequence performance, leading our authors to “therefore suggest that the pre-SMA activity during improvisation may be particularly related to decisions about timing and rhythmic patterning.” The PMD, on the other hand, has been implicated in the transformation of information held in ‘working memory’ into a sequence of movements in the correct order.
Thus, claim the authors, the role of the RPC in improvisation could be the timing (pre-SMA) and sequencing (PMD) of movements, such that the PMD is more involved with melodic structures and the pre-SMA with rhythmic structures.
Superior Temporal Gyri (STG)
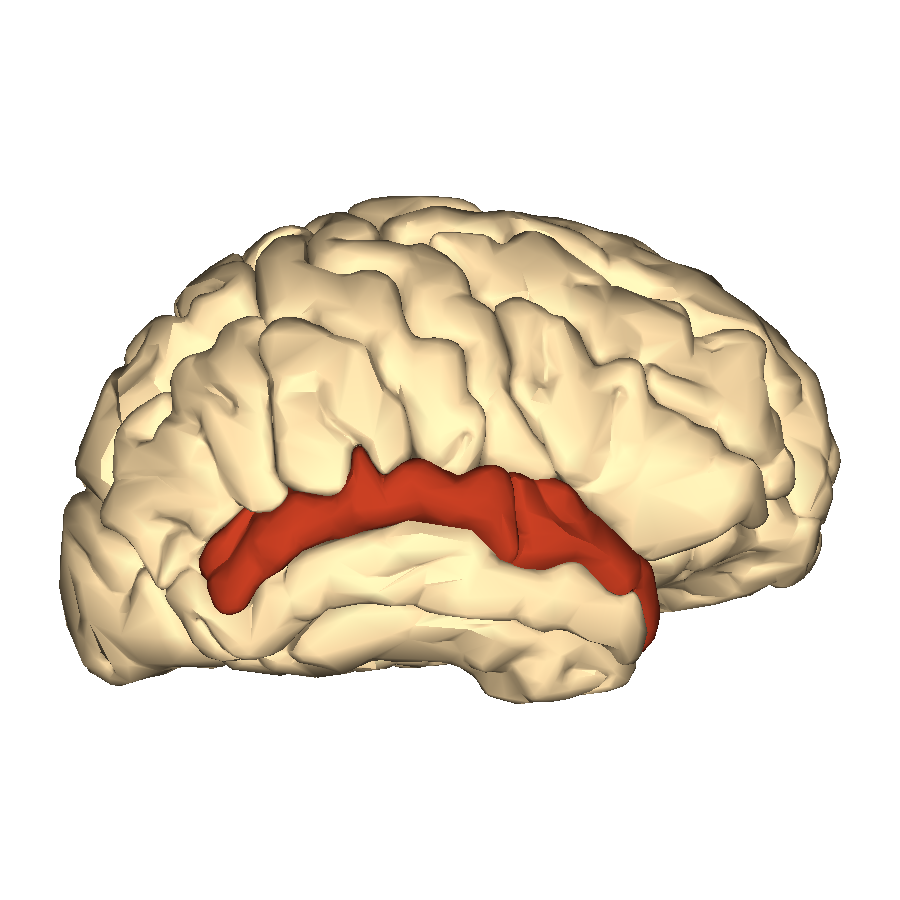
The posterior STG is a brain region that “has consistently been found active in studies that require auditory–motor integration, such as rhythmic sequence performance[…] and vocal rehearsal of words or music”. Bengtsson suggests that its increased activity evident in her pianists may reflect the increased reliance on “auditory-motor feedback loops” demanded by improvisation, “for example to adapt the improvisation to ongoing performance.”. The same region has also been shown to be involved in “auditory working memory”, and “thus may be of importance for auditory monitoring of the ongoing performance”. The posterior STG also has a role in word retrieval in linguistic tasks – perhaps, suggest the authors, it is similarly involved in the retrieval of grace notes, trills etc from a “common “vocabulary” of ornaments used in Western art music”
Middle occipital gyrus (MOG)
Heightened activity in this brain area, known to process higher-order visual input including reading music, is thought by the authors to “be due to a more intense visual processing of the musical score when this was used as a basis for improvisations, rather than as a template to recall a previous performance.”
In conclusion the authors declare “We have provided evidence that a set of frontal and temporal association areas [DLPFC, RPC and STG] are specifically involved in the free creation of musical structure during improvisation.”
Coda
Improvised Passacaglia in D minor
Alexander Weimann – harpsichord